Abychom byli schopni hovořit komplexně o hemoglobinu (Hb), je užitečné se nejprve zabývat myoglobinem (Mb), který je velmi podobný hemoglobinu, ale je mnohem jednodušší. Mezi hemoglobinem a myoglobinem existuje přísný vztah příbuznosti: obě jsou konjugovanými proteiny a jejich protetická skupina (non-proteinová část) je skupina hemu .
Myoglobin je globulární protein skládající se z jednoho řetězce asi sto padesáti aminokyselin (v závislosti na organismu) a jeho molekulová hmotnost je asi 18 Kd.
Jak je uvedeno výše, je opatřena hemovou skupinou, která je vložena do hydrofobní (nebo lipofilní) části proteinu, skládající se ze skládání připsatelných strukturám a-šroubovice vláknitých proteinů.
Myoglobin je složen převážně ze segmentů α-helixů přítomných v počtu osmi a sestává téměř výhradně z nepolárních zbytků (leucin, valin, methionin a fenylalanin), zatímco polární zbytky jsou prakticky nepřítomné (kyselina asparagová, kyselina glutamová, lysin a arginin); jedinými polárními zbytky jsou dva histidiny, které hrají zásadní roli v napadení kyslíku do skupiny hem.
Hemová skupina je chromoforová skupina (absorbuje se ve viditelné) a je funkční skupinou myoglobinu.
Viz také: glykovaný hemoglobin - hemoglobin v moči
Trochu chemie
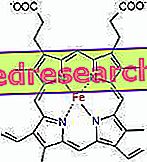
Spojení mezi protoporfyrinem a železem je typická vazba takzvaných koordinačních sloučenin, které jsou chemickými sloučeninami, ve kterých centrální atom (nebo ion) tvoří vazby s jinými chemickými druhy v počtu vyšším než je jeho oxidační číslo (elektrický náboj). V případě hemu jsou tyto vazby reverzibilní a slabé.
Koordinační číslo (počet koordinačních vazeb) železa je šest: kolem železa může být šest molekul, které sdílejí vazebné elektrony.
Aby se vytvořila koordinační sloučenina, trvá dva orbitály se správnou orientací: jeden je schopen "koupit" elektrony a druhý je může darovat
V hemu tvoří železo čtyři rovinné vazby se čtyřmi atomy dusíku, které jsou ve středu proto-porfyrinového kruhu a pátou vazbu s proximálním histidinovým dusíkem; železo má šesté volné koordinační spojení a může se vázat na kyslík.
Když je železo ve formě volného iontu, jeho d- orbitály mají stejnou energii; v myoglobinu je iont železa vázán na protoporfyrin a histidin: tyto druhy magneticky narušují orbitály železa; rozsah odchylky bude odlišný pro různé orbitály podle jejich prostorové orientace a rušivých druhů. Vzhledem k tomu, že celková energie orbitálů se musí vrátit na konstantní, rušení způsobuje energetickou separaci mezi různými orbitály: energie získaná některými orbitály je ekvivalentní energii ztracené ostatními.
Jestliže oddělení, které nastane mezi orbitals není příliš velké, vysoce-spinové elektronické uspořádání je preferováno: lepící elektrony pokoušejí se uspořádat sebe v paralelních otočkách v několika možných sub-úrovních (maximální multiplicity); na druhé straně, pokud je porucha velmi silná a existuje velká separace mezi orbitály, může být výhodnější spárovat vazebné elektrony v nižších energetických orbitálech (nízká rotace).
Když se železo váže na kyslík, molekula má nízkou spinovou dispozici, zatímco když má železo šesté volné koordinační spojení, molekula má vysokou rotaci.
Díky tomuto rozdílu ve spinu jsme díky spektrální analýze myoglobinu schopni pochopit, zda je vázaný kyslík (MbO2) nebo ne (Mb).
Myoglobin je protein typický pro svaly (ale nejen ve svalech).
Myoglobin se získává z velryby spermií, ve které je přítomen ve velkém množství a poté purifikován.
Cetaceans mají dýchání jako lidské bytosti: mají plíce, které musí absorbovat vzduch dýchacím procesem; velryba spermie musí nést co nejvíce kyslíku ve svalech, které jsou schopny akumulovat kyslík tím, že ho váží na přítomný myoglobin; kyslík je pak uvolňován pomalu, když je kytovník ponořen, protože jeho metabolismus vyžaduje kyslík: čím větší je množství kyslíku, který je spermie schopna absorbovat a čím více kyslíku je k dispozici během ponoru.

Aby bylo maso více či méně červené, je obsah hemoproteinů (to je hém, který dělá maso červené).
Hemoglobin má mnoho strukturních analogií s myoglobinem a je schopen vázat molekulární kyslík reverzibilně; ale zatímco myoglobin je omezen na svaly a periferní tkáně obecně, hemoglobin se nachází v erytrocytech nebo červených krvinkách (jsou to pseudo buňky, to znamená, že nejsou skutečnými buňkami), které tvoří 40% krve.
Hemoglobin je tetrametr, to znamená, že se skládá ze čtyř polypeptidových řetězců, z nichž každý má hémovou skupinu a dva po dvou (v lidské bytosti jsou dva alfa řetězce a dva beta řetězce).
Hlavní funkcí hemoglobinu je transport kyslíku ; další funkcí krve, ve které se hemoglobin podílí, je transport látek do tkání.
V cestě z plic (bohatých na kyslík) do tkání nese hemoglobin kyslík (současně se do tkání dostávají i jiné látky), zatímco v opačném směru s sebou nese odpad z tkání, zejména oxidu uhličitého produkovaného v metabolismu.
Ve vývoji lidské bytosti existují geny, které jsou exprimovány pouze po určitou dobu; proto máme různé hemoglobiny: fetální, embryonální, dospělého muže.
Řetězy, které tvoří tyto různé hemoglobins mají různé struktury, ale s podobností funkce, kterou vykonávají, je více méně stejná.
Vysvětlení přítomnosti několika různých řetězců je následující: během evolučního procesu organismů se také vyvinul hemoglobin, který se specializuje na přepravu kyslíku z oblastí, které jsou bohaté na nedostatečné oblasti. Na začátku evolučního řetězce hemoglobin přenášel kyslík v malých organismech; v průběhu evoluce organismy dosáhly větších rozměrů, proto se hemoglobin změnil tak, aby byl schopen transportovat kyslík do oblastí daleko od místa, kde byl bohatý; Za tímto účelem byly v průběhu evolučního procesu kodifikovány nové struktury řetězců tvořících hemoglobin.
Myoglobin váže kyslík i při mírném tlaku; v periferních tkáních je tlak (PO2) asi 30 mmHg: při tomto tlaku myoglobin neuvolňuje kyslík, takže by byl neúčinný jako nosič kyslíku. Hemoglobin má na druhé straně pružnější chování: váže kyslík při vysokých tlacích a uvolňuje ho, když tlak klesá.
Když je protein funkčně aktivní, může mírně změnit svůj tvar; například okysličený myoglobin má odlišnou formu od neokysličeného myoglobinu a tato mutace neovlivňuje sousední.
Diskurz je odlišný v případě asociovaných proteinů, jako je hemoglobin: když je řetězec okysličený, je indukován ke změně jeho tvaru, ale tato modifikace je tedy trojrozměrná, takže jsou ovlivněny i další řetězce tetrametru. Skutečnost, že řetězy jsou navzájem spojeny, vede k domněnce, že změna jednoho ovlivňuje i jiné sousední, i když v jiném rozsahu; když řetězec je okysličený, ostatní řetězce tetrametru vezmou “méně nepřátelský postoj” k kyslíku: obtíže se kterým řetězcové oxygenates klesá jak řetězy vedle toho okysličují sebe. Stejný argument platí pro deoxygenaci.
Kvartérní struktura deoxyhemoglobin je volána T (napjatá) forma zatímco to oxyhemoglobin je voláno R forma (povolený); v nataženém stavu existuje řada poměrně silných elektrostatických interakcí mezi kyselými aminokyselinami a bazickými aminokyselinami, které vedou k pevné struktuře deoxyhemoglobinu (to je důvod, proč "napjatá forma"), zatímco když je vázán kyslík, entita těchto interakce klesá (proto "uvolněná forma"). Kromě toho v nepřítomnosti kyslíku je náboj histidinu (viz struktura) stabilizován opačným nábojem kyseliny asparagové, zatímco v přítomnosti kyslíku existuje tendence proteinu ztrácet proton; to vše znamená, že okysličený hemoglobin je silnější kyselina než deoxygenovaná hemoglobie: bohr efekt .
V závislosti na pH se skupina hemu více či méně snadno váže na kyslík: v kyselém prostředí hemoglobin uvolňuje kyslík snadněji (napjatá forma je stabilní), zatímco v základním prostředí je vazba s kyslíkem silnější.

Každý hemoglobin uvolňuje 0, 7 protonů na příchozí mol kyslíku (O2).
Bohrův efekt umožňuje hemoglobinu zlepšit jeho schopnost přenášet kyslík.
Hemoglobin, který činí cestu z plic do tkání, se musí vyrovnat podle tlaku, pH a teploty.
Vidíme vliv teploty .
Teplota v plicních alveolech je asi o 1-1, 5 ° C nižší než venkovní teplota, zatímco ve svalech je teplota asi 36, 5-37 ° C; jak se teplota zvyšuje, klesá faktor nasycení (se stejným tlakem): to se děje proto, že se zvýší kinetická energie a disociace.

Existují i jiné faktory, které mohou ovlivnit schopnost hemoglobinu vázat se na kyslík, přičemž jedním z nich je koncentrace 2, 3 bisfosfoglycerátu .
2, 3 bisfosfoglycerát je metabolické činidlo přítomné v erytrocytech v koncentraci 4-5 mM (v žádné jiné části těla není přítomno v takové vysoké koncentraci).
Při fyziologickém pH je 2, 3 bisfosfoglycerát deprotonován a má na něj pět negativních nábojů; to jde do klín mezi dvěma beta řetězci hemoglobin protože tyto řetězce mají vysokou koncentraci pozitivních nábojů. Elektrostatické interakce mezi beta řetězci a bisfosfoglycerátovými řetězci poskytují systému určitou tuhost: získá se napjatá struktura, která má malou afinitu k kyslíku; během okysličování se vyloučí 2, 3 bisfosfoglycerát.
V erytrocytech je speciální přístroj, který převádí 1, 3 bisfosfoglycerát (produkovaný metabolismem) na 2, 3 bisfosfoglycerát tak, aby dosáhl koncentrace 4-5 mM, a proto je hemoglobin schopen výměny l \ t kyslíku ve tkáních.
Hemoglobin přicházející do tkáně je v uvolněném stavu (vázán na kyslík), ale v blízkosti tkáně je karboxylován a přechází do nataženého stavu: protein v tomto stavu má nižší tendenci vázat se kyslíkem, ve srovnání s uvolněným stavem proto hemoglobin uvolňuje kyslík do tkáně; kromě toho, v důsledku reakce mezi vodou a oxidem uhličitým, dochází k produkci iontů H +, a proto k dalšímu kyslíku v důsledku bohr efektu.
Oxid uhličitý difunduje do erytrocytů procházejících plazmatickou membránou; vzhledem k tomu, že erytrocyty tvoří přibližně 40% krve, bychom měli očekávat, že do nich vstupuje pouze 40% oxidu uhličitého, který difunduje z tkání, ve skutečnosti 90% oxidu uhličitého vstupuje do červených krvinek, protože obsahují enzym, který konvertuje oxidu uhličitého v kyselině uhličité vyplývá, že stacionární koncentrace oxidu uhličitého v erytrocytech je nízká, a proto je rychlost vstupu vysoká.

Dalším jevem, ke kterému dochází, když erytrocyt dosáhne tkáně, je následující: gradientem derivát HCO3- (oxid uhličitý) vychází z erytrocytů a pro vyvážení výstupu záporného náboje máme vstup chloridů, které určují zvýšení osmotického tlaku: k vyrovnání této odchylky je také vstup vody, který způsobuje otok erytrocytů (účinek HAMBURGER). K opačnému jevu dochází, když erytrocyt dosáhne plicních alveol: dochází k deflaci erytrocytů (účinek HALDANE). Žilní erytrocyty (přímo do plic) jsou tedy kulatější než arteriální.


